| VENTA IMAGENES FOTOS VIDEOS ALTA DEFINICION FAUNA FLORA HABITATS ACUATICOS RIOS VENEZUELA PIRANAS AMAZONAS FOTOGRAFIA VIDEOGRAFIA BAJO AGUA DULCE LAGOS SUBACUATICA FOTOGRAFOS VENEZOLANOS NATURALEZA |

Preservación del agua, flora, fauna y del medio ambiente en Venezuela
Impacto del Control de los Ríos |
Bol. Acad. C. Fís., Mat. y Nat. Vol. LXVIV No. 4 |
EL CONTROL DE LOS RÍOS Y SU IMPACTO SOBRE LA ICTIOFAUNA CONTINENTAL: UNA REVISIÓN
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
RESUMEN El incremento en la población humana y su estrecha dependencia del recurso hídrico, han influido cada vez más en la masiva construcción de obras de control de ríos. Estas son realizadas en el eje longitudinal o transversal del canal y alteran diferentes componentes del ecosistema lótico. Los cambios ambientales producidos tienen efectos de diferente magnitud e intensidad sobre las comunidades asociadas y la respuesta de las especies depende, entre otras, de características particulares de su historia natural. En el presente trabajo se señalan características e impactos de estas obras sobre las comunidades de peces, con énfasis especial en la construcción de embalses. La información revela impactos producidos por la alteración de la dinámica del flujo y cambios en la heterogeneidad espacial; de estas condiciones se derivan una serie de factores que afectan la estructura de la comunidad íctica y se discuten efectos sobre algunos aspectos ecológicos como la alimentación y reproducción de las especies. Se resalta además, el efecto negativo sobre las rutas migratorias de muchas especies y la escasa eficiencia de las medidas de mitigación y control de este efecto.
Palabras clave: peces, ecosistema lótico, embalses, impactos
Las comunidades acuáticas, entre ellas la íctica, adaptadas a las condiciones del ecosistema fluvial preexistente, responden en formas diferentes a los efectos generados por el control del río. Estas diferencias son influenciadas por el tipo de obra de control, su escala, los componentes de la dinámica fluvial que altere y principalmente por la historia de vida de las especies que conforman la comunidad. La evaluación del impacto de las obras de control de ríos, ha sido en muchos aspectos subjetiva como consecuencia del desconocimiento de las condiciones previas del ecosistema y las comunidades asociadas. Sin embargo, los impactos que se han identificado sobre la ictiofauna señalan en forma general una alteración negativa en la estructura de las comunidades asociadas y aunque algunas especies se ven favorecidas por las nuevas condiciones, muchas veces este bienestar es temporal e inestable. En el presente trabajo se recopila información concerniente a las obras de control de los ríos y su impacto sobre la ictiofauna continental, señalando el efecto diferencial según aspectos biológicos y ecológicos de las especies y algunas medidas de mitigación que han sido implementadas. Dentro de las obras de control de las aguas, se da un énfasis especial a la construcción de embalses, ya que es una de las prácticas más comunes e importantes dadas las necesidades humanas de suministro seguro de agua y energía, y posiblemente, una de las de mayor impacto sobre las comunidades de peces.
RESULTADOS
Los principales factores que se controlan son los componentes del caudal del río: velocidad y volumen de agua y sus efectos sobre el canal y área circundante como la erosión del lecho y los márgenes, la sedimentación y las inundaciones. Según el objetivo o el componente del caudal que se requiera controlar, las obras de control de los ríos pueden efectuarse en el perfil longitudinal o en el transversal del río. Obras Longitudinales: como su nombre lo indica, son obras marginales realizadas en el perfil longitudinal del río y tienen como objetivos principales: el control de la erosión en los márgenes, inundaciones, encauzamiento hacia una estructura de paso (un puente o una alcantarilla) y la desviación del canal. Estas obras poseen extensiones variables y su construcción es más común en tramos del río que recorren zonas urbanas. – Obras Marginales de Encauzamiento: buscan encauzar una corriente natural hacia una estructura de paso como un puente, alcantarilla, etc. Poseen transición de entrada y salida, y tener diferentes dimensiones. – Obras de Protección de Márgenes: son muros o revestimientos resistentes a la fuerza del agua que fluye por el canal, y principalmente evitan la erosión de los márgenes. El revestimiento es realizado con diferentes materiales: rocas, plástico, corazas metálicas, llantas (cauchos) y gaviones, estos últimos quizá los más comunes consisten en una estructura metálica en forma de escalera enrejada llena de rocas. Obras Transversales: se construyen de ribera a ribera del río (eje transversal) y su magnitud y estructura dependen del objetivo del control. En algunos casos se controla el caudal en un pequeño tramo del río, utilizando materiales convencionales como sacos de arena, escalas de roca, etc., esta regulación puede dirigirse a la protección de estructuras de paso, suministro de agua en zonas agropecuarias e incluso el control de inundaciones. Dentro de las obras transversales, la de mayor magnitud e importancia, respecto a impactos sobre la ictiofauna, es la construcción de embalses. Los embalses son cuerpos de agua artificiales señalados como híbridos entre río y lago, por su naturaleza intermedia entre lótica y léntica. Son construidos represando un río o corriente, o por la construcción de paredes en un sitio impermeable con un canal riverino como suplemento de agua (Ramírez, 2000). Los principales objetivos de la construcción de un embalse son: a) Regular el suministro de agua a la agricultura en lugares de pluviosidad fluctuante o irregular o donde la lluvia cae fuera de la estación favorable;
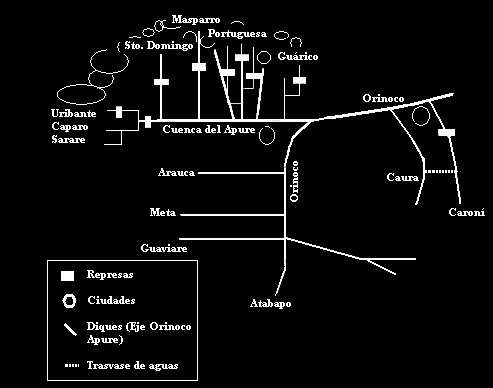
b) Hacer regables y así aprovechar terrenos que se encuentran muy alejados de la red hidrográfica principal, o incluso pertenecientes a otras cuencas; c) Obtener energía eléctrica; y d) Suministrar agua a ciudades o industrias. Gracias a la utilidad de estos cuerpos de agua, su construcción se ha hecho generalizada en varios países donde la demanda energética e hídrica se ha incrementado significativamente. En los Estados Unidos entre 1930 y 1975 se construyeron alrededor de 10000 embalses y esta actividad no se ha detenido (Taylor et al. 2001); en Venezuela, la construcción de estos cuerpos de agua artificiales que comenzó intensivamente en los años 60, se ha incrementado significativamente en las últimas décadas (Cressa et al. 1993; Machado-Allison, 2005) y el número de embalses de alta capacidad existentes está alrededor de 100 (Páez et al. 2001). En Colombia, el área embalsada alcanza las 63.238 ha, con una capacidad total de 8684 millones de metros cúbicos. Estas cifras son casi el triple del área de los principales lagos o lagunas de Colombia, la cual se estima en 19.031 ha. Estos reservorios se encuentran ubicados en los pisos térmicos fríos (23%), en los cálidos (47%) y en las altitudes medias (30%) (Valderrama, 1995). En estos y otros países, la mayorí parte del área embalsada se encuentra en zonas donde está asentada gran parte de la población y por consiguiente donde se establece el eje de mayor desarrollo productivo, como es el caso de la cuenca del Orinoco en Venezuela, donde casi todos los ríos de la cuenca provenientes de los Andes se encuentran afectados (Machado-Allison, 2005) (Figura 1), o la cuenca del río Magdalena en Colombia, que drena de sur a norte el territorio nacional, y recibe la influencia de algunos de los más grandes asentamientos urbanos (Galvis y Mojica, 2007). Los embalses están por tanto localizados sobre cuencas alteradas y de gran importancia socioeconómica y como consecuencia, están profundamente modificados, bien sea por procesos acelerados de colmatación y eutrofización, por la llegada de materiales alóctonos provenientes de la cuenca, o por materiales autóctonos provenientes del metabolismo interno (Ramírez, 2000). Dentro de las características básicas de los embalses se encuentran (Ramírez, 2000; Marquez y Guillot, 2001): * Las cubetas de los embalses son alargadas y su línea de costa muy irregular y desarrollada, dependiendo de la geomorfología del valle inundado. * Son de flujo más lento que el de los ríos y más rápido que el de un lago. * Los sedimentos de fondo son de origen alóctono. El material autóctono se sedimenta en las partes lacustres del embalse y el material alóctono se acumula cerca de la zona fluvial. * Las fluctuaciones de nivel son más intensas, frecuentes e irregulares que en los lagos. Mientras estos suelen desaguar por la superficie, la descarga en embalses es de aguas intermedias o profundas. * Las fluctuaciones de nivel se dan en periodos cortos y la tasa de renovación es más elevada que en los lagos. * La biomasa es menor y posee una tasa de renovación más elevada. * Los embalses nacen en condición heterotrófica, y posteriormente se presenta el proceso de digestión de la materia orgánica que se hallaba en el fondo del terreno inundado. * Las características geológicas de la cuenca son determinantes en aspectos como la físico química de las aguas y la carga de sedimentos, las cuales a su vez tienen influencia sobre la productividad biológica y características tróficas del embalse. * La naturaleza de la cubeta del embalse, la vegetación y el tipo de suelos inundados, constituyen un factor determinante del estado trófico del embalse y sus características ecológicas. * La termoclina que puede llegar a establecerse en los embalses suele ser más pequeña y a veces más profunda que en lagos comparables.
El viento que en los embalses sopla generalmente en el sentido de su eje longitudinal, puede tener mayor influencia en la irregularidad de la estratificación térmica.
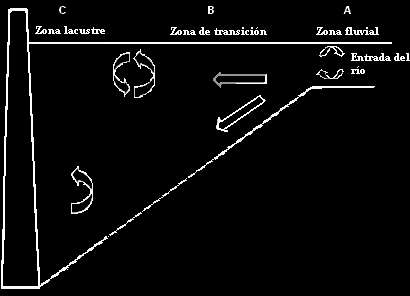
Zonificación de los Embalses: la construcción de un embalse representa una alteración del ecosistema fluvial original e implica la transformación de la naturaleza lótica del río a una principalmente léntica del embalse. El carácter gradual y permanente de este cambio una vez establecido el embalse le confiere diferentes características, donde según Thornton (1990) pueden distinguirse tres zonas principales en un perfil longitudinal: zona fluvial o cola del embalse, zona de transición y zona de presa (Figura 2).
ARRIBA: Figura 1. Esquema gráfico de la ubicación de la mayoría de los embalses, relación con ciudades,
ARRIBA: Figura 2. Zonación de un embalse. Modificado de Thornton et al. 1990.
A) Zona Fluvial o Cola del Embalse: corresponde a la unión entre el afluente y las aguas del embalse. En esta área se presenta una disminución de la velocidad del flujo mientras aumenta la tasa de sedimentación de partículas de arena y limo gruesas. Por diferencia de densidad, el afluente se hunde al entrar en el embalse, ya que las aguas del río tienden a estar algunos grados por debajo de las del embalse, dejando en la superficie los materiales flotantes (macrófitas, troncos, basuras, etc.). Esta es una zona generalmente poco profunda y bien mezclada; sin embargo, la elevada turbidez del agua y la disminución en la velocidad del flujo disminuye la actividad fotosintética. B) Zona de Transición o Media del Embalse: en esta zona se combinan características del río y del lago. La cuenca del embalse se profundiza y amplía, haciéndose más similar a un lago; el flujo de agua es más reducido y muchos materiales en suspensión se depositan, permitiendo mayor penetración de la luz que a su vez favorece un incremento en la productividad planctónica. El incremento en la penetración de la luz puede ser gradual o abrupto dependiendo del flujo de agua. Aquí se sedimentan arcillas y limos de tamaño grueso a medio y materia orgánica particulada. C) Zona Lacustre o Presa del Embalse: en esta zona, la más profunda del embalse, la naturaleza léntica se acentúa y las características ambientales son muy similares a las de un lago. La penetración de la luz aumenta y por consiguiente la producción fitoplanctónica. La columna de agua exhibe una estratificación más definida, y existe una limitación potencial de la producción por escasez de nutrientes, dado que la tendencia es hacia condiciones oligotróficas.
2. Impacto del control de los ríos sobre la ictiofauna continental. La naturaleza y la intensidad de los impactos generados por las obras de control de ríos, dependen de la historia de vida de la ictiofauna local. Las especies perciben y responden al cambio ambiental según características como estrategias reproductivas, patrones de migración, especializaciones tróficas y el grado de “preadaptación” a ambientes lacustres o lénticos, así como de las características particulares de la obra construida como su magnitud, localización, morfología, hidrología, diseño, procedimientos operacionales, entre otras (Tabla 1) (Agostinho et al. 2007; Machado-Allison, 2005). En el caso de la construcción de embalses, la World Commission of Dams (WCD 2000) propone tres categorías para los impactos de esta obra a nivel de ecosistemas. * Primer orden: incluyen consecuencias físicas, químicas y geomorfológicas producto del bloqueo del río y de alteraciones en la distribución espacio-temporal de la cubeta del embalse. * Segundo orden: involucran cambios en la productividad primaria y en la estructura del canal, comprendiendo el tramo represado y principalmente el segmento delante de la barrera. * Tercer orden: incluyen las modificaciones en las comunidades de invertebrados y peces producto de los impactos de primer y segundo orden. Una vez establecida la obra, las nuevas condiciones pueden ser restrictivas para algunas especies pero favorables para otras, y las respuestas son diferenciales también respecto a la fase de construcción. Las características del ecosistema lótico que se ven principalmente afectadas por las obras de control son las derivadas de la heterogeneidad espacial y la dinámica del flujo de agua: a) Heterogeneidad Espacial: la heterogeneidad ambiental le confiere estructura espacial a las comunidades bióticas que allí habitan. Las diferencias en tipos de sustrato determinan el establecimiento de diferentes comunidades animales y vegetales bentónicas en el ecosistema. Estas comunidades dependen entre sí como sustrato, recurso alimenticio y refugio. Una vez reducida la heterogeneidad espacial, puede modificarse la dinámica comunitaria que garantiza la coexistencia de especies competidoras, de presas y depredadores y alterarse así la diversidad de la comunidad. Las riberas de los ríos están conformadas generalmente por vegetación leñosa que constituye un bosque de galería, fuente importante de materiales alóctonos, que al entrar al agua incrementan la heterogeneidad espacial y los recursos (espaciales y alimenticios), los cuales son aprovechados por una alta diversidad de organismos acuáticos (insectos, crustáceos, peces y otros organismos). Además, la constante erosión que ejerce el agua sobre los márgenes, descubre las raíces de los árboles y retira trozos de sedimento creando cuevas, las cuales son utilizadas como refugio o nido durante el proceso reproductivo por un gran número de especies ícticas (Machado-Allison 1990, 1992, 1994 y 2005). Las obras marginales de encauzamiento y de protección de márgenes reducen significativa-mente este componente del ecosistema lótico mediante la homogenización del tipo y forma del sustrato afectando el ciclo vital de estos organismos. b) Dinámica del Flujo de Agua: se ve afectada principalmente por las obras de control transversal, aunque las de tipo longitudinal pueden alterarla también a pequeña escala aumentado la velocidad del flujo debido a una reducción en la erosión de materiales del lecho y los márgenes. En las obras transversales, las que más afectan la dinámica del flujo de agua son los embalses. En estos cuerpos de agua las fluctuaciones de nivel son mucho más intensas, frecuentes e irregulares que en los lagos y son inducidas por el tipo de operación hidráulica. Mientras los lagos suelen desaguar por la superficie, la descarga en embalses es de aguas intermedias o profundas. Los embalses tienen fluctuaciones grandes de nivel en períodos cortos y tasas de renovación más
Tabla 1. Esquema mostrando actividades, acciones, efectos e impactos sobre la comunidad acuática con el cierre de un caño: Un ejemplo. (Fuente: Machado-Allison, 2005).
elevadas que en los lagos. Al igual que en los lagos, la forma y orientación de la cubeta respecto al régimen dominante de vientos es determinante en la circulación de las aguas en los embalses y de sus características de estratificación y mezcla. Embalses alargados, sobre todo si lo son en la dirección del viento dominante, se mezclan con mayor frecuencia y de manera más uniforme que embalses de formas redondeadas o con muchas bahías y recodos. En el trópico, donde la temperatura de las aguas tiende a ser estable y no se propicia la mezcla por diferencias de densidad, estos factores mencionados son cruciales en los procesos de estratificación y mezcla. Los patrones que se establecen en estos procesos son decisivos en la determinación de la naturaleza ecológica del embalse. Los embalses oligomícticos (escasa mezcla) tienden a conservar condiciones eutróficas, mientras los polimícticos (bien mezclados) tienden a ser holomícticos (mezcla en toda la columna de agua) y evolucionan hacia condiciones mesotróficas (Marquez y Guillot, 2001). La edad o etapa de evolución de la obra de control, en el caso de obras transversales como los embalses, influencia condiciones distintas en el ecosistema acuático y las comunidades asociadas. Para los embalses neotropicales, Agostinho et al. (1999) resaltan diferentes eventos desde la construcción hasta el envejecimiento de estos cuerpos de agua (Tabla 2). La construcción de un embalse tiene efectos de diferente índole sobre varios aspectos biológicos y ecológicos de la ictiofauna, a nivel poblacional y comunitario, algunos de ellos se describen a continuación con base en información proveniente de diferentes estudios realizados en zonas tropicales y templadas.
3. Efectos sobre el ciclo reproductivo. Una clasificación de los peces está relacionada con la habilidad de completar su ciclo de vida dependiendo del acceso o no a ambientes lóticos. De esta forma se distinguen las especies riverinas obligatorias, las cuales desovan y completan su ciclo solamente en el corredor o canal principal del río y las especies riverinas facultativas que pueden realizar partes de su ciclo de vida tanto en aguas corrientes como lacustres o de sabanas o bosques inundables (Holden, 1979). Se ha reconocido que estos dos grupos de peces reaccionan en formas distintas a la construcción de embalses. Casi todas las especies riverinas obligatorias son afectadas con el represamiento y con consecuencias que van desde reducciones poblacionales hasta la extinción local de muchas de ellas (Penczak et al. 1998). La principal causa para esta afectación es la irrupción del continuo del río (Ward y Stanford, 1983). Ya que la construcción del embalse trae consigo cambios en el régimen de flujo, cantidad y composición de sólidos suspendidos, alternancia de parámetros físicos y químicos del agua y reducción en número de microhábitats, se da un bloqueo de las rutas de migración, cambios en el tipo y abundancia de alimento, número y distribución de zonas de cría y por consiguiente la disminución en el reclutamiento de las poblaciones (Welcomme, 1985). Las especies riverinas facultativas en contraste, son generalmente las más resistentes a alternancias en el flujo de agua y el tamaño y forma del canal del río y en algunos casos, sus densidades se ven afectadas positivamente con este tipo de cambios en el hábitat. En estas especies se han registrado cambios positivos en su densidad poblacional y factor de condición (grado de bienestar obtenido de la relación longitud-peso) en las primeras etapas de establecimiento de embalses. Sin embargo, no es posible generalizar el carácter positivo de la respuesta de estas especies al ambiente embalsado. Kruk y Penczak (2003), abordaron este tema en un estudio realizado aguas abajo y aguas arriba de un embalse localizado en el curso medio del Río Warta en Polonia y sus resultados mostraron que no solo las especies riverinas obligadas sufrieron por el efecto del embalse. Aspectos particulares de la biología de las especies facultativas influyeron significativamente en su respuesta al represamiento. Anguilla anguilla, Lota lota, Silurus glanis y Esox lucius disminuyeron notablemente en número y frecuencia de aparición, incluyendo su aparente ausencia en algunos años después del represamiento. Mientras las otras especies involucradas en el estudio, Stizostedion lucioperca, Gymnocephalus cernuus, Perca fluviatilis, Rutilus rutilus, Abramis brama y Blicca bjoerkna, presentaron incrementos de decenas y cientos, hasta miles por hectárea. Se destaca el papel que cumplen aspectos de la historia de vida de estas especies, ya que pueden generar respuestas diferenciales, siendo la más importante para ese estudio, la flexibilidad en el sitio de desove y la alimentación, ya que fueron las especies con tendencia a la especialización en la dieta y con requerimientos particulares de sitios de desove las que presentaron reducciones en su densidad poblacional. En el Valle del Río Magdalena, Galvis y Mojica (2007) señalan un efecto negativo del embalse de Betania sobre especies de Salminus e Ichthyolephas, que comparten áreas de desove con especies de tierras bajas, pero viven en los rápidos y corrientes de las montañas en altitudes por debajo de 1000 m de altitud.
Tabla 2. Principales eventos en el ecosistema acuático durante la evolución de un embalse
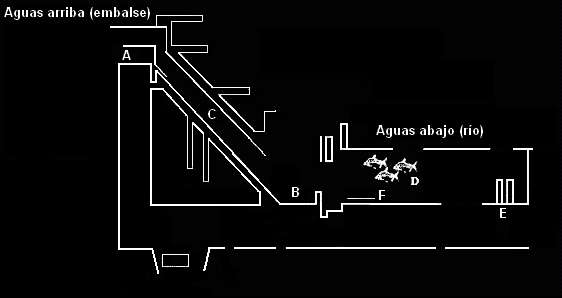
Por otro lado, en Venezuela la construcción de represas ha permitido la proliferación de especies “plaga” como es el caso del caribe colorado (Pygocentrus cariba) en represas como la del Guárico (estado Guárico) y Las Majaguas (estado Portuguesa). Además, se han impedido las “ribazones” (migraciones con fines reproductivos) de varias especies de importancia comercial como las cachamas (Colossoma macropomum), los coporos (Prochilodus mariae) en los llanos y el suapire (Mylesinus schomburgki) en el Caroní (Machado-Allison, 2005). El impacto sobre las especies migratorias es particularmente obvio. Normalmente, las zonas de reproducción se encuentran aguas arriba de las zonas de cría y alimentación. Su estrategia reproductiva involucra la utilización de la corriente y el desborde de las aguas como mecanismos de dispersión de huevos y larvas. Las zonas marginales inundables constituyen las principales áreas de cría por sus condiciones adecuadas de protección y disponibilidad de alimento (Wootton, 1990). La construcción de represas interfiere de varias formas en esta dinámica migratoria. Sin embargo, el grado de afectación es diferente según la ubicación de la presa en relación con el circuito migratorio, distinguiéndose las poblaciones cuyo circuito migratorio es interrumpido por la presa y las poblaciones en las que este queda incluido en el sistema formado por el embalse y el tramo del río aguas arriba, o se desarrolla totalmente en el tramo aguas abajo de la presa (Espinach y Ríos 1997). En el caso de las poblaciones de peces migratorios con área de reproducción aguas arriba de la presa y áreas de cría y alimentación aguas abajo, los impactos más evidentes son: 1. Bloqueo de las migraciones reproductivas. Este efecto se ha buscado mitigar con el funcionamiento de instalaciones para el paso de los peces (esclusas, escaleras, ascensores, etc.) (Figura 2). 2. En la ruta migratoria, muchos peces se enfrentan al paso a través de turbinas y vertederos que aumentan la mortalidad de adultos en migración de retorno, así como de juveniles y larvas. 3. Las modificaciones en el régimen de flujo aguas arriba y aguas abajo de la presa generan alteración del comportamiento migratorio. 4. La extensión y ubicación de las áreas usadas por los peces para el desove y la cría de larvas y juveniles se ven alteradas por la inundación. El flujo de agua, aunque reducido, es fundamental en estas áreas. Puede que este tipo de ambientes persistan aguas arriba del embalse y sus afluentes, sin embargo, la ubicación a mayor distancia y la menor velocidad de deriva, pueden impedir la llegada de larvas y juveniles, cuya habilidad para realizar este recorrido depende de condiciones ambientales como la disponibilidad de alimento y la intensidad de la depredación. Las especies con rutas migratorias localizadas aguas arriba de la presa, no sufren la interrupción física, pero sí pueden verse afectadas por: 1. Diferente grado de distanciamiento de las zonas idóneas para la reproducción. 2. Reducción de la velocidad de deriva. 3. Reducción en superficie y número de las áreas de cría, que repercute además en el aumento de la intensidad predatoria en estas áreas. Las poblaciones aguas abajo de la presa, estarían afectadas por: 1. La ausencia de la periodicidad natural en el flujo producida por la operación de la central y el vertedero. 2. Alteración del comportamiento reproductivo y migratorio. 3. Disminución y desaparición de áreas aptas de cría de larvas y juveniles (áreas inundadas p.e.). 4. Reducción en la abundancia y diversidad en la oferta de alimento proveniente de deriva fluvial o material orgánico alóctono. Muchas de estas especies migratorias alcanzan grandes tallas y son de importancia en la explotación pesquera comercial, artesanal y deportiva. Sin embargo, estas pesquerías en embalses han presentado reducciones importantes que se han atribuido a un grupo de factores como: la sobrepesca, la ocupación y uso incorrecto de las tierras circundantes, y especialmente al represamiento de los ríos (Agostinho et al. 2005). En ausencia de medidas de mitigación del efecto del embalse sobre las rutas migratorias, el grado de impacto causado por la estructura del mismo dependerá de la habilidad y posibilidades del pez de desovar en la vecindad inmediata del embalse, o de buscar áreas alternativas o remanentes aguas abajo o arriba. Sin embargo, existen aún muchas dudas sobre la flexibilidad de las especies migratorias en sus requerimientos espaciales para completar el ciclo reproductivo (Ribeiro et al. 2007). Diferentes estudios al respecto, han revelado la complejidad de los patrones migratorios en las especies de peces neotropicales. Ribeiro et al. (2007) realizaron un estudio en el embalse Porto Primavera del Río Paraná usando técnicas de marcaje y recaptura y encontraron cierta flexibilidad en el comportamiento reproductivo, ya que sus resultados indicaron que por lo menos una parte del grupo que migra es capaz de cambiar su ruta migratoria después de encontrar un obstáculo. Este comportamiento es fundamental, ya que puede asegurar la reproducción y por ende la permanencia de las poblaciones, aún en situaciones en las cuales las principales rutas de migración se han perdido. Otros estudios soportan esta tendencia, por ejemplo en Prochilodus argenteus se encontró que puede usar efectivamente tributarios aguas abajo del embalse Tres Marías (Río San Francisco-Brasil) para la reproducción (Sato et al. 2005). Sin embargo, es importante aclarar que el hecho de que las especies puedan encontrar rutas alternas, esto no indica la efectividad del desove y el reclutamiento, ya que si los peces se concentran por una cantidad prolongada de tiempo, la demora en desovar puede resultar en la reabsorción de los oocitos maduros (Agostinho et al. 1993). Se ha detectado además que algunas especies migratorias, como Prochilodus lineatus y Pterodoras granulosus, tienen la capacidad de detectar y evitar las áreas represadas, ya que las “reconocen” como poco favorables para el desove y se estimula una mayor velocidad de migración aguas arriba (Agostinho et al. 1994). En el estudio de Ribeiro et al. (2007), se capturaron individuos junto a la presa de contención, algunos fueron liberados aguas arriba y otros fueron mantenidos aguas abajo del embalse. Más del 50% de los peces recapturados aguas arriba, habían sido liberados aguas arriba; esto señala, que existe un porcentaje importante de peces que después de ser transferidos, no proceden con su migración aguas arriba. Los autores sugieren que estos resultados indican como los peces pueden verse desorientados al llegar al ambiente léntico y comenzar a migrar en direcciones distintas. Esta situación tiene implicaciones directas en la efectividad de las estructuras de paso y elevadores que han sido considerados como medidas de mitigación para el efecto de los embalses sobre estas especies, y muestra además que muchos peces dejados aguas arriba de la presa se regresan pasando por turbinas y vertederos que ponen en riesgo su integridad física. Por otro lado, Espinach y Ríos (1997), evaluaron diversos impactos del represamiento sobre la ictiofauna en el embalse de Salto Grande en Uruguay en diferentes períodos a partir de la construcción. El efecto encontrado sobre las especies migratorias fue diferencial. La mayoría de ellas experimentaron una reducción en su abundancia en el año siguiente al embalsado y posteriormente, algunas mostraron evidencias de adaptación a las nuevas condiciones, como es el caso de Leporinus obtusidens y Salminus brasiliensis. Otras en cambio, como Sorubim lima, Pseudoplatystoma coruscans y Oxydoras kneri continuaron la tendencia declinante hasta prácticamente desaparecer de las capturas. En Colombia, Galvis y Mojica (2007), afirman que la construcción de hidroeléctricas en la cuenca del Río Magdalena, no ha generado reducción en sus pesquerías, debido a que estos embalses han sido construidos muy por encima de los límites superiores de la migración de los peces. Sin embargo, en Venezuela varias represas se han construido aguas abajo en las vías migratorias de especies de peces (p.e. coporos, cachamas, palometas, suapires, etc.) de importancia comercial como es el caso del represamiento de los ríos Portuguesa, Masparro, Boconó, Guárico y Caroní. Por otro lado, este represamiento ha causado cambios en la dinámica del flujo hídrico, pérdida de la capacidad del agua de entrar a las sabanas, esteros y bosques durante el período de lluvias y eliminar potenciales áreas “nursery” para la reproducción y desarrollo de larvas y juveniles de organismos acuáticos (Machado-Allison, 2005), ver más adelante. 4. Impacto sobre la estructura de la comunidad de peces La construcción de un embalse provoca serios cambios en el ambiente y una pérdida relevante de biodiversidad con consecuencias impredecibles a largo plazo en el medio (Britsky, 1994). La evaluación e interpretación de los cambios temporales en términos de la composición de especies, su abundancia, sus estrategias tróficas y reproductivas permiten evaluar la calidad del ambiente donde viven y así sus requerimientos de hábitat, para inferir sobre sus posibles respuestas al cambio ambiental (Petesse et al. 2007). En este aspecto se ha obtenido la siguiente información: 1. Los embalses deben ser colonizados por los peces de los ríos represados; el cambio del ambiente lótico al léntico no es asimilado por la mayoría de las especies y solo unas pocas, por lo común las habitantes de los remansos del río, logran adaptarse al embalse. Esto determina un cambio en las proporciones de ocurrencia de ciertas especies y una redistribución espacial y temporal de las especies, que modifican los principales atributos de la estructura comunitaria: composición, riqueza y diversidad (Marquez y Guillot, 2001). Sin embargo, no es posible hablar de una única comunidad nueva establecida, ya que desde la inundación inicial y las siguientes etapas de evolución del embalse, las condiciones ambientales se modifican y estructuran de diferente forma la comunidad. Muchas veces la alteración inicial de la comunidad no se puede identificar, debido al desconocimiento de la comunidad habitante del río y sus afluentes antes de su represamiento. 2. La identificación de las especies presentes en el embalse y el estudio de las variaciones en su abundancia relativa a lo largo del tiempo, son necesarios para inferir las tendencias en la transformación de las comunidades de peces, detectar los efectos de posibles perturbaciones, formular medidas de protección en caso necesario, y realizar el seguimiento de las consecuencias de su aplicación (Espinach y Ríos, 1997). 3. El embalse, puede generar fragmentación de hábitat al actuar como barrera física pero también como barrera ambiental debido a la calidad del agua, impidiendo el establecimiento de algunas especies. En el estudio de la literatura se encontraron algunos ejemplos que documentan los factores anteriormente indicados. Así, en el sur de Brasil, existe un complejo lagunar costero formado por las lagunas de Patos y Mirim, las cuales se encuentran comunicadas por un canal natural llamado San Gonzalo. En 1977, se construyó un embalse en este canal para evitar la entrada de agua salada desde la laguna de Patos hacia Mirim, ya que de esta última se extrae agua dulce para riego de cultivos de arroz. Antes de esta construcción, se había registrado la ocurrencia común de peces marinos y estuarinos tales como: Micropogonias furnieri, Mugil platanus y Paralichthys orbigmyanus que servían de soporte importante para la pesca artesanal de la zona. Burns et al. (2006), estudiaron los efectos del embalse sobre las comunidades de peces en estas dos lagunas. Los resultados indican que el embalse actúa como una barrera que impide la entrada de agua salada hacia la laguna de Mirim y este es un factor abiótico que muestra una distribución espacial discontinua que coincide con la encontrada para la ictiofauna. Esto soporta la hipótesis de que los embalses construidos cercanos a las lagunas litorales pueden alterar el hábitat y la estructura de la comunidad, interrumpiendo sus conexiones naturales entre sistemas acuáticos adyacentes, tales como el marino, estuarino y dulceacuícola. Por otro lado, en el estudio del impacto de la Hidroeléctrica Samuel sobre las comunidades de peces del río Jamari en Brasil, el número total de especies en las fases pre y post-represamiento fue el mismo; sin embargo, se observó una acentuada alteración en la composición de la ictiofauna producto de la formación del embalse (Santos, 1995). Al evaluar los dos períodos, el autor concluye que las comunidades de peces del canal principal del río se vieron más afectadas que aquellas que habitaban los lagos marginales; según sus resultados es posible identificar además la redistribución espacial de algunas especies en las diferentes zonas del embalse muestreadas, y la aparente desaparición de otras, entre ellas aquellas que emprenden una o dos migraciones anuales a lo largo del curso del río, aún cuando antes del represamiento fueran comunes de lagunas adyacentes o remansos del río. En otro caso, en un pequeño embalse de los Estados Unidos al sur de Illinois en el Kinkaid Creek, se comparó la comunidad de peces presente antes y después del represamiento. La riqueza de especies fue casi la misma, 38 y 39 especies antes y después respectivamente, sin embargo, la composición fluctuó significativamente. 13 de las 38 especies colectadas antes del represamiento no fueron encontradas después y 13 de las 39 encontradas después no habían sido colectadas antes de la construcción. Pasó de ser una comunidad dominada por la familia Cyprinidae a ser dominada por Centrarchidae (54.4%). Se registró la extinción local de seis especies con requerimientos de flujo permanente de agua. Los autores reportan como una causa primordial de estas extinciones y modificaciones en la estructura comunitaria, la descontrolada introducción de especies con fines de pesca comercial y deportiva (Taylor et al. 2001). Otra de las causas del cambio en las proporciones de abundancia entre las especies de la comunidad íctica es la mortalidad producida por la calidad del agua en el embalse, que ha sido registrada en muchos cuerpos de agua. En algunos casos, el fenómeno se ha relacionado con vientos fuertes que rompen la estratificación térmica y transfieren, desde el hipo hacia el epilimno, anhídrido sulfuroso, muy tóxico para los peces. Otra de las causas es la anoxia nocturna, que puede ser causada por la proliferación de algas (blooms), como por ejemplo, de la especie Botryococcus braunii, o de cianofíceas tóxicas. En todos los casos, las altas mortalidades parecen estar relacionadas con la eutroficación como causa básica; esta se agudiza por la estratificación térmica que propicia la anoxia hipolimnética, fenómeno que además puede generar muerte de organismos por agua vertida del embalse hacia el efluente (Márquez y Guillot, 2001). Además, el desconocimiento de la fauna íctica nativa que habita el o los ríos a represar y el interés de realizar una explotación pesquera en el embalse, ha dado lugar a la introducción de especies exóticas, que en muchos casos gracias a la preadaptación a ambientes lénticos, logran establecerse con éxito en este ecosistema y desplazar o incluso conducir a la extinción local de especies nativas, por interacciones como la competencia y la depredación principalmente. Finalmente, es necesario indicar que el estudio de la composición y abundancia de las especies en la comunidad íctica del embalse son fundamentales en la formulación de planes de aprovechamiento piscícola. Sin embargo, a pesar de que en el embalse habiten especies de importancia comercial, el aprovechamiento en cultivos puede verse afectado por condiciones físicas, químicas y biológicas de cuerpo de agua. Un ejemplo lo constituye el embalse Lagartijo en Venezuela, sistema poco favorable para el cultivo de peces, dadas las bruscas y pronunciadas variaciones de nivel a las que se ve sometido durante el año en lapsos muy cortos de tiempo, y además por la mala calidad el agua bombeada desde el río Tuy, la cual contiene cloro y elementos producto de la contaminación industrial fundamentalmente (Infante et al., 1993). Algunos cambios en la composición de la comunidad también pueden estar influenciados por el hábito alimenticio de las especies de peces y relacionados por tanto con la oferta de recurso en el nuevo ambiente léntico. Este aspecto será discutido a continuación. 5. Impacto según los hábitos alimenticios: No todas las especies que habitan el río logran adaptarse al embalse, esto determina que nichos y hábitats importantes queden sin ocuparse y que en consecuencia la eficiencia total del ecosistema en la utilización de energía disponible sea inferior a lo posible. Por ejemplo, las especies planctófagas especializadas son escasas en áreas neotropicales continentales, ya que en los ríos el plancton es un recurso limitado en contraste con su gran abundancia en los embalses; dada esta condición, gran parte del plancton se transfiere a los sedimentos, donde se acumula y solo la menor parte, es utilizada por vías detríticas y aprovechada indirectamente por los peces, entre los cuales se cuentan los comedores de fondo (bentófagos) bien sea omnívoros o detritívoros (Marquez y Guillot, 2001); como es el caso de Eremophilus mutisii, una especie de Trichomycteridae, de hábitos bentófagos, que incluye en su dieta el consumo masivo de cladóceros y copépodos en el área embalsada, mientras en la zona lótica, se alimenta principalmente de macroinvertebrados acuáticos (Andrade, 2006a). En esta especie se registró además una reducción importante en la talla, peso y factor de condición promedio a lo largo del gradiente ríoembalse en una zona alto Andina de Colombia (Andrade, 2006b). En algunos embalses de Venezuela, de los que se dispone de información sobre los hábitos alimenticios de la fauna íctica y el zooplancton, este constituye un recurso importante en la dieta de 13 especies de peces (larvas, alevines y especies pequeñas) (López et al., 2001) como por ejemplo Roeboides dayi (Ortaz et al., 1999; Ortaz et al., 2003). La importancia de los peces zooplanctófagos en embalses se relaciona también con la posibilidad de aplicar técnicas de biomanipulación de la trama trófica. Esto funciona como una medida complementaria para la reducción de las abundancias del fitoplancton en aquellos ambientes eutróficos (Ortaz et al. 2006). Para lo cual es ideal estudiar los hábitos alimenticios de las especies nativas que habitan el embalse, evitando al máximo la introducción de especies, ya que en muchos casos esto puede traer cambios en la composición de especies sobre la estructura trófica del ambiente y repercutir en la calidad del agua (Agostinho et al., 1999). Muchos cambios en la estructura de la comunidad de peces después del represamiento del río pueden ser explicados por los hábitos alimenticios de las especies. Por ejemplo, los Curimatidae presentan hábitos bentónicos y se constituyen como un grupo de peces típico de ambientes lacustres o remansos de río, su recurso alimenticio está conformado por sedimentos y material orgánico particulado, el cual se deposita en el fondo después del represamiento como consecuencia tanto de la carga transportada por las aguas como por la alta producción biológica autóctona; en este grupo se han registrado aumentos importantes en la abundancia de las especies en ambientes embalsados (Santos 1995, Espinach & Ríos 1997). Como fue evidente en los dos primeros años posteriores al llenado del embalse de Salto Grande en el Río Uruguay, las especies de las familias Prochilodontidae, Curimatidae y Loricariidae, presentaron un aumento significativo en su abundancia, debido probablemente a la gran cantidad de alimento disponible en forma de materia orgánica degradable proveniente de los ecosistemas terrestres inundados. Posterior a este lapso, se registró una disminución en las densidades poblacionales de estos grupos, hasta que en el período a partir de los nueve años posteriores a la inundación nuevamente aumentaron, posiblemente favorecidas por el incremento en la superficie y la sedimentación de detritos orgánicos en el embalse. En el caso de Prochilodus lineatus, el aumento fue posibilitado además por el reemplazo de las poblaciones afectadas por el bloqueo de sus migraciones, por otras cuyas rutas migratorias quedaron incluidas en el embalse y el tramo aguas arriba, anteriormente poco numerosas. En este embalse se registró también el aumento en la densidad poblacional de especies asociadas a ambientes pelágicos o de aguas libres que se alimentan en forma exclusiva u oportunista de elementos del plancton, como el bagre Parapimelodus valenciennis, como era previsible teniendo en cuenta la expansión y desarrollo del ambiente pelágico al conformarse el embalse, donde inicia además la proliferación de las comunidades planctónicas. Una situación similar se observó en la Hidroeléctrica Samuel, donde los peces planctófagos como Hypophthalmus edentatus y Schizodon fasciatus, antes raros en el río Jamari, proliferaron significativamente en el embalse en función del incremento de sus fuentes de alimento: el plancton, las macrófitas acuáticas y las algas asociadas. El aumento de estas poblaciones, que corresponden a presas potenciales, favoreció además la proliferación de especies depredadoras como Serrasalmus rhombeus y Cichla monoculus (Santos 1995). Por otro lado, en el embalse de Salto Grande (Río Uruguay), no todas las especies depredadoras se vieron favorecidas. Mientras Acestrorhynchus altus y Cyrtocharax squamosus aumentaron significativamente, la guabina Hoplias malabaricus mostró una reducción muy marcada, registrando en los primeros años del represamiento capturas de 10Kg/Batería/Noche (Batería: 900 m2 de redes de ahorque) y posteriormente apenas 0.65 Kg/Bat/Noche. Esto puede explicarse posiblemente por los efectos negativos de las fluctuaciones de nivel del embalse sobre esta especie, que deposita sus huevos en nidos ubicados en aguas poco profundas. Se registró además la desaparición de Pyaractus mesopotamicus y Brycon orbigyianus, especies con régimen alimenticio fundamentalmente frugívoro, como producto posiblemente de la deforestación de la vegetación autóctona marginal del río Uruguay (Espinach y Ríos, 1997). En general en los embalses el recurso natural responsable del mantenimiento de la productividad es principalmente de origen alóctono. La ictiocenosis parece depender en gran medida de la cadena detritívora. La vegetación circundante y los pulsos estacionales de inundación son los principales proveedores de alimento a la fauna íctica (Alvim y Peret, 2004). 6. Obras para la mitigación de impactos sobre la comunidad de peces A pesar de los múltiples impactos ocasionados por la construcción de embalses, los esfuerzos para la mitigación de los mismos se han concentrado en el bloqueo de las rutas migratorias. Muchos de estos mecanismos están basados en escaleras, estructuras que reducen la velocidad del agua y la pendiente, de tal forma que los peces pueden ascender y pasar el embalse (Agostinho et al., 2007). Otra herramienta son las esclusas para peces (Figura 3), las cuales basadas en diferencias de caudal atraen a los peces hacia un compartimento, luego los transportan hacia arriba y los liberan en el embalse. Las esclusas consisten en un conducto inclinado que vincula al río aguas abajo con el embalse. En cada extremo del conducto se encuentra una cámara provista de compuertas, la inferior, o cámara de acumulación y la superior o de salida. Aguas abajo de la cámara inferior se extiende una pileta en la que se encuentran ubicadas las entradas para los peces. Estos ingresan a la pileta atraídos por el flujo de agua proveniente del sistema, debido a que los peces en su migración se orientan por la corriente. El criterio más común para medir la eficiencia de estos mecanismos es el número de peces que ascienden. Sin embargo, en muchos casos se ha encontrado cierta selectividad de la estructura respecto a su eficiencia dependiendo de la especie. En las esclusas del embalse de Salto Grande en Uruguay, Espinach y Ríos (1997), encontraron que la cantidad y composición específica de los peces transportados por la estructura varió en función del caudal erogado durante la etapa de llamada hacia la compuerta de entrada, así como de la hora del día. Siluriformes y Gymnotiformes fueron comunes durante la noche, mientras otros grupos como los Clupeiformes mostraron valores máximos de captura durante el día. Esto sugiere la necesidad de estudios cuidadosos del manejo de estas estructuras, ya que durante el día se requieren diferentes caudales para transportar diferentes especies. El problema en muchos casos es que se considera a estas estructuras como el fin del efecto y no como lo que son en realidad, herramientas para el diseño de políticas de manejo que deben ser monitoreadas (Agostinho et al. 2002). En la situación ideal, estas estructuras deberían tener baja selectividad de especies, permitiendo el mantenimiento de los movimientos de los peces, llevando a una estructura comunitaria similar a la observada antes del represamiento del río, con la misma proporción de especies aguas abajo.
Figura 3. Corte de la esclusa tipo Borland del Embalse de Salto Grande (Uruguay). A. Cámara superior o de salida. B. Cámara inferior. C. Ducto inclinado. D. Pileta de acumulación de peces. E. Entrada a la pileta. F. Orificios de comunicación entre la pileta y la zona de descarga de turbinas.
Entre los aspectos más críticos para la eficiencia de estas estructuras están el mecanismo que atrae a los peces, el cual debe permitir a los cardúmenes reconocer su entrada rápida y exactamente. Si el pez no reconoce la entrada, puede permanecer en la vecindad por un período muy largo, demorando la migración y poniendo en peligro la reproducción, o resultando en el no ascenso de la escalera. Las condiciones físicas y químicas del agua como la temperatura, la velocidad del agua y el oxígeno disuelto aguas abajo del embalse, donde los cardúmenes están migrando río arriba generalmente se acumulan, son frecuentemente estresantes para los peces y pueden llevar a la reabsorción de los productos gonadales por parte de las hembras (Agostinho et al. 1993). Una esclusa para peces representa una pérdida de agua para la generación de energía y su funcionamiento puede entrar en conflicto con el aprovechamiento hidroeléctrico del embalse, especialmente en regímenes de descarga limitados. Este conflicto es mitigado ya que afortunadamente las migraciones ocurren durante la época de lluvias. Los peces muestran diferentes preferencias y habilidades con respecto al flujo de agua, el cual puede presentar una descarga adecuada para un grupo de peces pero desfavorable para otros. La dinámica de las corrientes río abajo del embalse, relacionada con la distribución de los cardúmenes de peces en este tramo del río, limita las alternativas para colocar la entrada de las esclusas o escaleras. La relación de esta dinámica con los procedimientos operacionales de un embalse y las limitaciones impuestas por la entrada de agua en el éxito de la entrada de peces a la esclusa o la escalera son factores adicionales que complican el funcionamiento de estas medidas de mitigación. En adición a la atracción, la efectividad de los pasos para peces depende de la velocidad del agua, la turbulencia, temperatura, turbidez y las grandes burbujas de agua a lo largo de su longitud (Rodríguez et al., 2006). Después de que el pez entra, debe ser capaz de superar estos obstáculos y alcanzar el embalse, lo cual depende de las diferentes habilidades de las especies para pasar corrientes, y esto frecuentemente constituye una fuente de selectividad en esta etapa del viaje. Esta selectividad es un cuello de botella importante para las iniciativas de transportar peces con el objetivo de conservar sus poblaciones y la biodiversidad. Esto sin embargo no es lo único; la intensificación de la depredación, la demora en el desplazamiento hacia los sitios de cría, el regreso de los migrantes y el reclutamiento de nuevos individuos a las poblaciones aguas abajo del embalse, son otros impedimentos que pueden resultar en un paso para peces ineficiente o incluso ambientalmente indeseable (Agostinho et al. 2007). Las evidentes diferencias en la respuesta de las especies de peces a las modificaciones del ambiente lótico, hacen suponer que el estudio de los efectos de las obras de control de ríos sobre la ictiofauna, debe ser coherente con la complejidad de estas comunidades y la historia natural de cada una de las especies que las conforman, para de esta manera lograr evaluar objetivamente el impacto y formular acciones eficientes en la mitigación de efectos, entendiendo que el aprovechamiento del recurso hídrico y la conservación de la ictiofauna no deben verse como objetivos divergentes, sino más bien como herramientas en la sostenibilidad del uso de los recursos. El Futuro: Una reflexión final. Dada la incertidumbre actual, los planes de desarrollo en países latinoamericanos y la poca o nula atención que se le ha prestado a esta actividad en nuestro país es necesario tomar en cuenta ciertas condiciones. Petts (1990) nos ilustra sobre aquellas necesarias para lograr una regulación sostenible en nuestros ríos: 1) Viendo hacia el futuro, es inocente pensar que un manejo científico adecuado puede prevenir la pérdida de algunas partes de los componentes de un ecosistema. Los ecosistemas naturales son sistemas dinámicos de corto, mediano y largo plazo y este dinamismo incluye componentes azarosos importantes. Sin embargo, la ciencia debe tener un papel importante en este problema. El incremento del esfuerzo e investigaciones científicas dedicadas a la interfase de hidrología, geomorfología y ecología, para entender la vulnerabilidad de los ecosistemas y el avance de las bases científicas para regulaciones o normativas segundarias, son necesarias y urgentes, por lo tanto la integración de equipos multi o transdisciplinarios es de gran importancia; 2) Un desarrollo sostenible requiere del cambio en el comportamiento humano acerca de de cómo se percibe el mundo. Es todavía popular la creencia de que el agua representa un recurso natural capaz de desarrollar energía (p.e. electricidad) a un costo (económico o ambiental) mucho más bajo que el petróleo o energía nuclear. Esta percepción está basada en cuatro argumentos: a) agua dulce natural corriendo hacia los océanos es agua perdida; b) los humedales o pantanos son tierras pobres y representan un peligro sanitario; c) las inundaciones son una amenaza inaceptable para las tierras (agrícolas) y la vida humana; y d) la producción de hidroelectricidad es una opción ecológicamente sostenible para el desarrollo de programas de energía. Claramente como ha sido manifestado por numerosos autores (ver: Machado-Allison, 2005; Petts, 1990), esta visión debe ser cambiada mediante procesos educativos a todos los niveles indicando entre otras cosas, que los humedales o pantanos permanentes o temporales poseen un valor inmenso en el mantenimiento de las poblaciones de organismos acuáticos, incluyendo aquellos que frecuentemente participan en la dieta de la fauna silvestre y de poblaciones humanas. Que las inundaciones son necesarias para completar ciclos biológicos y biogeoquímicos; y que la producción de hidroelectricidad no es automáticamente una opción ecológicamente sostenible y en muchos casos ha sido un factor de perturbación y desaparición de numerosos ecosistemas en nuestro país; 3) Los desarrollos exitosos del uso de nuestros recursos naturales deben estar basados no solamente en avances tecnológicos científicos y sociológicos, sino también en una buena coordinación y administración efectiva del agua, su descarga y calidad y el manejo adecuado de las tierras. Esperamos, que el desarrollo de los planes de incremento de la capacidad de agua almacenada en nuestros países, de ahora en adelante tome en cuenta el pasivo ambiental producido y en estos planes sea incorporado el componente ambiental para lograr el menor daño posible, un manejo adecuado y la conservación de nuestros recursos naturales.
LITERATURA CITADA |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IR A: ARTICULOS SOBRE AGUA DULCE | IR A: AGUA DULCE |
| N E W |
TODAS LAS IMAGENES Y VIDEOS EN AQUATIC-EXPERTS ESTAN DISPONIBLES PARA LA VENTA EN: WWW.MIKOLJI.COM |